Abstract
Cajal, the “father” of neurobiology, used Golgi’s silver stain to visualize neurons, which he represented as extended, arborized cells with indirect, synaptic contacts to one another (1900–1914). But he represented the neuron as floating in space (“naked neuron”). By contrast, Golgi claimed a Perineural Net (PNN) around the neuron, which Cajal dismissed as a “staining artifact”. Notwithstanding, modern analytic and microscope techniques revealed an Extracellular Matrix (nECM) around the neurons, through which non-synaptic signals could pass.
Cajal also enunciated 4 principles of neural signaling. The neurobiologist Hebb [1] interpreted these as “Synaptic Plasticity” (SP). He ascribed the basis of learning and memory to the increased number and functionality of synaptic contacts. Subsequently, Arshavsky [2] accused Hebb of 7 sins”, of failing to address many issues critical to modeling neural memory. We note that Hebb and following generations of neurobiologists continued Cajal’s “original sin”, of ignoring the implications of neural shape, thereby overlooking the presence of nECM.
As unction to redeem these sins, we offer a tripartite mechanism whereby cognitive units of information (cuinfo) are encoded as metal-centered complexes within the nECM, the “memory material” around the neurons. Neurotransmitters (NTs) permit the “chemo-coding” of emotive states, not available to any other coding scheme (Baudot, Braille, binary, trinary, Morse, electronic).
One can no longer evade the inadequacies of the Cajal/Hebb model of exclusive synaptic signaling, which require a rethinking the canons of neurobiology. The novel tripartite mechanism, augments the concept of “synaptic plasticity” and provides a chemo-dynamic model of neural coding of memory.
Keywords
Cognitive information, Metal complexes, Emotions, Neurotransmitters
Background
“Memory is a mystery as deep as any that psychology can propound.”
– William Bateson
“It is obvious that nerve impulse is somehow converted into thought, and that thought can be converted into nerve impulse. And yet, all this throws no light on this strange conversion.”
– Roger Penfield
Modern neurobiologists posit that memory results from the cumulative performance of sets of synaptically connected neurons, predicated on the neural model first described by Cajal [1–10]. The terms “Synaptic Plasticity” (SP), “Long Term Potentiation” (LTP), “connectivity” and the like, are currently used to describe the ability to recall. However, such terms lack biochemical definition and do not suggest a coding system. To regain focus, we reexamine the origins of neurobiology.
History
Essentially, Cajal used Golgi’s silver stain method to visualize neurons. Cajal saw and drew the neuron as an arborized cell with indirect, synaptic contacts to others. But the neuron’s exquisite shape was presented as if it were floating in space, with nothing surrounding it (“naked neuron”). By contrast, Golgi claimed a Perineural Net (PNN) around the neuron, which Cajal dismissed as a “staining artifact”.
Technical aside: The chemistry of silver salt underlies its utility as a stain for neurons. It is based on the affinity of soluble Ag+ for the hydrophobic lipid bilayer of the neural membrane, where it is reduced to insoluble Ag, which oxidizes to form insoluble black Ag2O [11, 12]. The stain thus reveals neural membrane shape at high b&w definition, effectively a photograph.
Silver Staining Equation
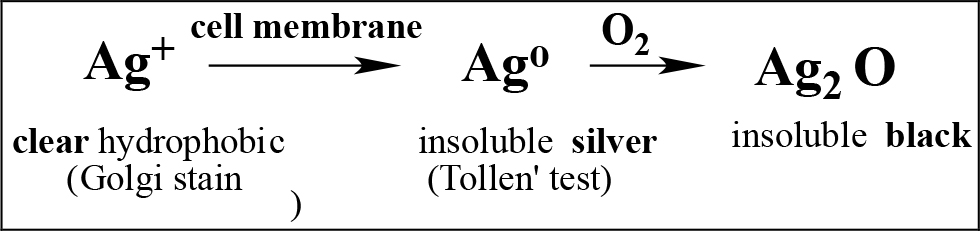
But as the silver stain does not react with polysaccharides of the nECM or most proteins, it did not reveal the nECM web enshrouding the neuron, which remained invisible and unconsidered.
Chemistry aside, Cajal did not infer the reality of the nECM from neural morphology. He did not recognize that the neural shape itself “spoke” about the cell’s intimate contact with its surroundings
(Figure 1A). It was as if he were a gardner who wanted to understand plant biology, but ignored the soil around the roots or the air around the leaves of the plant (Figure 1B).
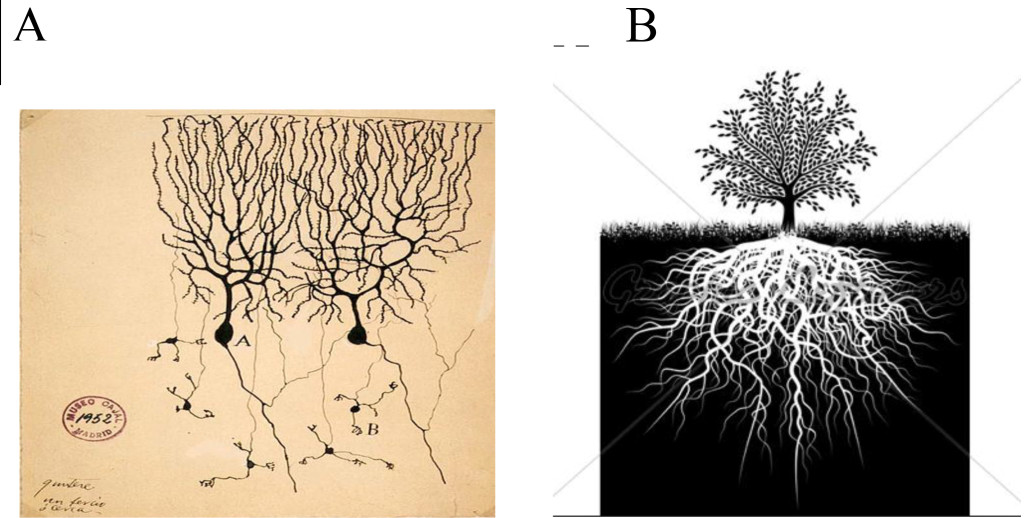
Figure 1. Reverse/ transpose panels A to B and B to A.
A. Cajal drawing (circa ~1911) of a neural net with dendrites extending into the surrounding area, many with no synaptic contact. The nECM is ignored (as “background”), with no function relating to signaling between neurons; hence “naked neurons”. B. Contrast-image
Consequently, Cajal ruled out non-synaptic signaling through the nECM. Based on his vision of neural connectivity, he proposed 4 principles of neural signaling.
Cajal’s 4 Principles [13]
- The neuron is the elementary signaling unit of the nervous system.
- The axon of the neuron communicates with other neurons only at specialized, non-contact regions, gaps called “synapses”.
- A given neuron will only signal with some specific cells but not with others.
- A neural signal travels in only one direction.
A schematic of Cajal’s idea of a neural net is provided in
Figure 2 A.
Based on Cajal’s principles, McCullogh and Pitts [14] mathematically described sets of synaptically connected neurons, uni-directionally signaling in binary modes (Equation 1).
Equation 1:
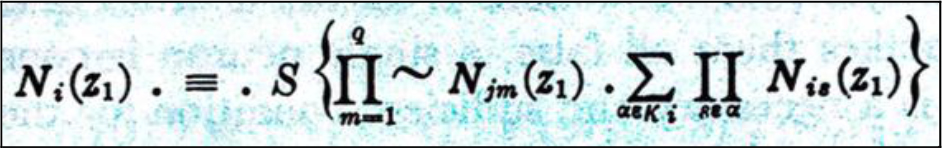
Pioneers of the “Information age”, von Neumann, Shanon and Schroedinger, attended McCullogh’s lecture at the 1948 Hixon Symposium [14, 15]. Ironically, this mathematical approach helped establish the theory and practice of electronic microprocessor memory at the core of modern computer chips. Though impressive, the equations did not throw much light on biological neuron mentation.
Continuing in the “Cajalian” vein, the neurobiologist Hebb [1] also ignored the nECM. He formulated a theory of “Synaptic Plasticity” (SP) wherein the basis of learning and memory was due to the increased number and functionality of neural synaptic contacts. An example of Hebb’s mathematical approach (Equation 2), reads:
Equation 2:
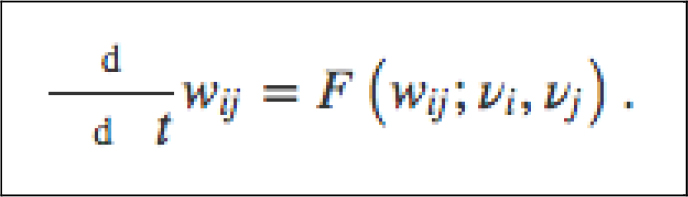
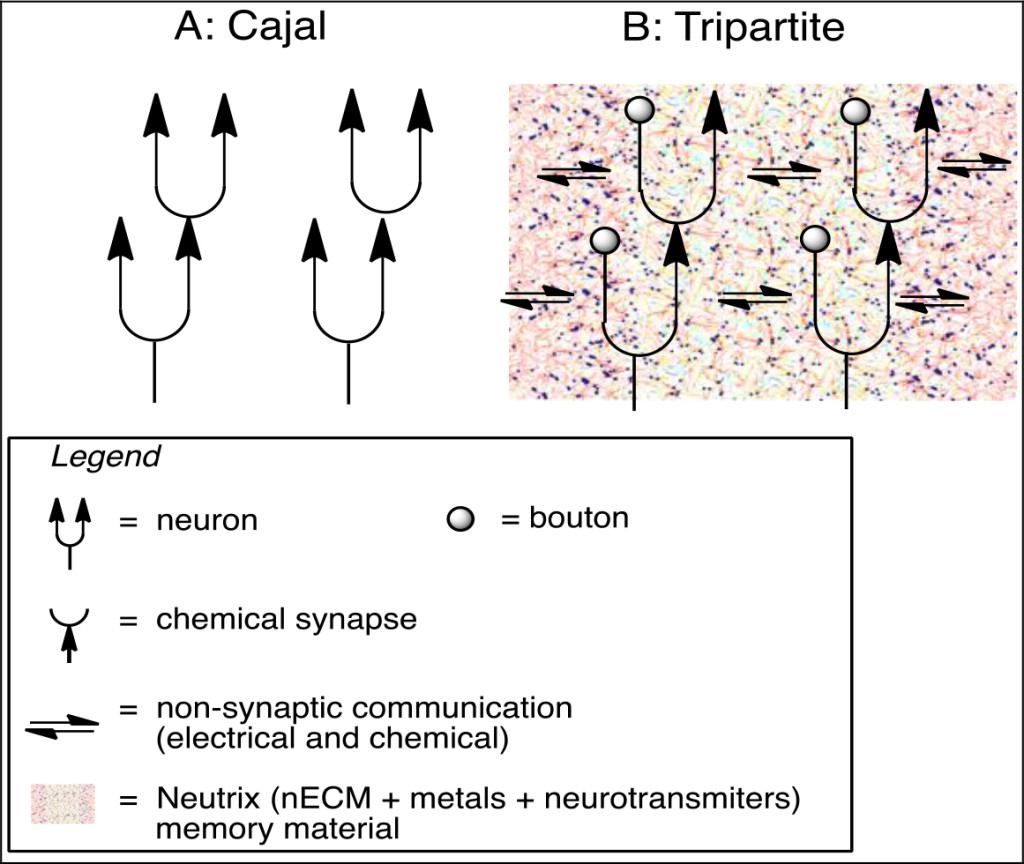
Figure 2. A: Schematic of Cajal’s neural net composed of 4 cells in synaptic contact with one another. Note that the neural environment is ignored i.e. “naked” neurons. B. A corrected tripartite schematic of a neural net, enmeshed in a surrounding “neutrix” ( nECM + metals and neurotransmitters (NTs)), engaged in non-synaptic, as well as synaptic signaling.
The “Hebbian” model of memory [1, 15–20] ascribed to the following precepts:
- Memory is represented by the joint activation of (sparse) groups of synaptically connected neurons.
- Learning results from the strengthening (increased function) of neural synaptic connections, termed plasticity or Long Term Potentiation (LTP).
“Mechanisms of learning and memory reside not in the special properties of the neuron itself, but in the connections it receives and makes with the neural net.”
– Kandel [12]
But saying that synaptic connection between neurons are “strengthened” or “tagged” [18] does not describe the molecular details whereby they encode and store persistent memory in accessible form.
Consider the reality of the brain’s neural nets. Since the early 1960’s and onward,
Golgi’s PNN, now called nECM, was rediscovered, characterized by analytic techniques and visualized by Scanning and Transmission Electron Microscopy (SEM and TEM respectively) [20–26]. Today, all neurobiologists admit that neurons are encased in a 3D matrix. Thus, the reality of the nECM has been recognized, but not yet internalized as having functional significance for learning or memory.
Reservations have been raised about the Cajal/Hebb model of neural signaling [2, 26–31]. In particular, it was noted that there are non-synaptic signaling pathways through the ubiquitous extracellular matrix (nECM) around all neurons [32–34]. Still today, most neurobiologists attempt to correlate learning and memory simply with changes of synapse number and functionality, termed Long-Term Potentiation (LTP), and do not account for the non-synaptic dendrites.
Hebb’s 7 “Sins”
In the light of the inadequacies of the LTP model, Arashavsky accused Hebb of 7 “sins” [3], of failing to address many issues critical to modeling neural memory.
- The synaptic plasticity hypothesis cannot explain the long-life persistence of memory.
- The suggestion, that the same mechanism operates for memory storage and recall, is seriously flawed
- Memory acquisition and storage have different localizations.
- ‘‘Synaptic’’ and ‘‘system’’ memory consolidations have different temporal characteristics.
- Reconsolidation of memory is not ‘‘predicted by traditional theories of memory consolidation”. Persistent declarative memory, stored in the brain through structural modifications in synaptic connections, “is incompatible with the phenomenon of memory reconsolidation after retrieval”.
- Neurogenesis occurs in the adult brain. Replacing old neurons with new neurons which still retain memory is puzzling; something basic in LPT must be missing.
- The synaptic plasticity hypothesis does not explain the specific memory impairments present in Alzheimer’s disease.
We note other failings:
- The neuron should be described as a polyvalent electro-chemical cell, not a binary (on/off) electrical devise.
- Mathematical descriptions of neural code cannot encode emotions, the basis for mentation.
Doctrinal Guidelines
It is generally accepted that neural mental processing is governed by the laws of chemistry and rules of biology. In today’s dogma of neurobiology, Synaptic Plasticity (SP) enshrines the ideas of Cajal and Hebb and many other neurobiologists [35, 36]. But it is hard to devise a synaptic connectivity code that would persist beyond a few seconds and provide emotive context.
One asks: What can a scientist refer to when advocating a mechanism for coding an emotive event experienced by a neural net?
Just like “information”, “cognitive information” requires a physical embodiment to achieve persistence [36], not simply a dynamic connection between neurons. What is the physicality of the memory trace, the engram?
As we grope for enlightenment, we realize that we require a specific language to comprehend the linkage between the physiology of our bodies and the psychic talents of our brains. To that end, we do not enlist the equations of mathematics [38, 39] or the algorithms of the computer model, but the concepts and iconography of chemistry [40, 41], which has been successfully used to clarify many other, previously mysterious aspects of our biological being, like metabolism, breathing, (i.e. Krebs cycle, hemoglobin) [39], blood coagulation (i.e. cascades of Factors) [42, 43] and reproduction (i.e. DNA → RNA → protein) [44].
What are the doctrinal guidelines that a scientist can refer to when advocating a mechanism for a psychic state experienced by the neural net of any creature?
The 7 characteristics and traits that one needs to address include:
- Process: A credible encoding mechanism for neural memory based on generally accepted biochemical principles, with components available to neurons in an aqueous milieu.
- Kinetics: Molecular-scale encoding/decoding process, faster than the rate of neural firing (<100 ms).
- Capacity: Large storage capacity for physically encoding cog-info.
- Energy: Low energy requirements (<400 cal/day).
- Storage: of cog-info for short and long durations.
- Loss: Forgetting as a loss of memory code.
- Universality: Applicable to all animals with neural circuitry.
Tripartite Mechanism of Memory
We propose that the neurons employ their surrounding nECM (Figure 1B) as a “memory material”. The diffusible metals and Neurotransmitters (NTs) perform as dopants to encode cognitive information (cog-info), at select addresses within the nECM to form “cognitive units of information” (cuinfo, (singular/ plural), metal-centered complexes represented as chemographic icons (Figure 3) detailed by Marx & Gilon [45, 46].
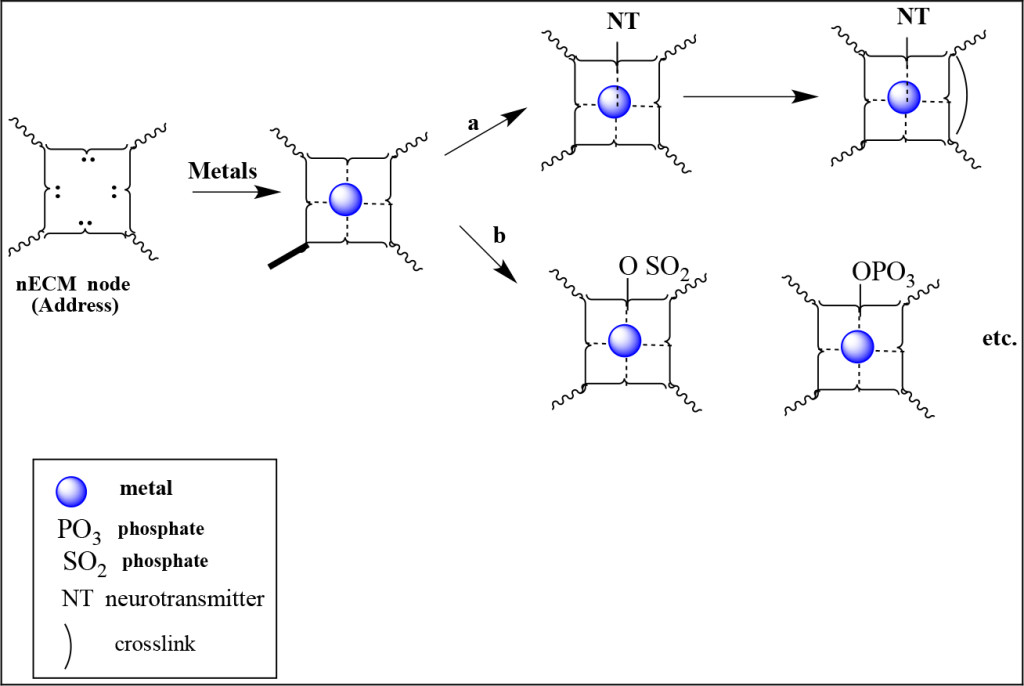
Figure 3. Chemographic representation of the encoding of cog-info. An nECM address can react with a metal resulting in a cuinfo, which can be tagged. The cuinfo can form a ternary complex by binding NT to the metal, sunsequently crosslinked (route a), or the cuinfo can be derivatized by chemical reactions (route b). Both pathways add unique encoding tags. The NTs add psychic dimensions to the cuinfo; the cross-links ensure stability.
Monovalent metals form relatively unstable complexes; polyvalent metals are generally more stable. Some could also engage in redox (Fenton) reactions, with attendant covalent modifications involving new condensation or cross-linking reactions. Thus, these reactions provide the neuron with a large encoding repertoire. Such a system was contemplated by Fodor [47], but not detailed. We present a chemographic shorthand in Figure 3.
Feelings, Emotions and Memory
The terms “feeling” and “emotion” are often used interchangeably [46]. However, we employ them as distinct terms referring to different physiologic reactions and psychic states:
- “Feelings” (often considered psychically [48]), actually relate to body sensations (light, sound, pain, balance, hunger, thirst, etc.) generated by specifc sensors to outside stimuli, which are accompanied by body reactions and corresponding psychic states. They are mediated with biologic modulators, called neurotransmitters (NTs) (Table 1) [49–52]. Astrocytes also release neuroactive molecules (gliotransmitters) to modulate neural signaling [52, 53].
The “meaning” of any stimulus set in memory, is based on its immediate “sensate value”, established by NTs. Any sensation (mild or acute) is “felt” physiologically and psychically, concomitant with the release of NTs during neural signaling (Table 1).
Table 1. Neurotransmitters (NTs), which effect both Physiologic reactions and Psychic States
|
Neurotransmitter (NT) |
Physiologic Reactions* |
Psychic Effects! |
|
Biogenic amines (8) Amino acids (>10) Neuropeptides (>70) Acetylcholine (1) NO (1) Endocannabinoids (>10) |
Breathing Blinking Blood Pressure Cold Contraction Muscles Coughing Crying Dilation of Muscles Dilation of Pupil Drooling Erection Evacuation Fever Goose Bumps Heart Beat Heat Hunger Pain Seeing Smell Thirst Touch |
Anxiety Aggression Awareness Depression Dreams Fear Hate Joy Love Paranoia Sadness Sex Drive Sociability |
*No Memory required ! Emotions require memory
- “Emotions” are remembered “feelings”, “chemo-coded” with neurotransmitters (NTs), as represented in Figure 3 and detailed elsewhere [45, 46]. NTs can attach to a cuinfo via a metal complexation bond and endow it with emotive (subjective) quality.
Accordingly, “feelings” do not require memory, whereas “emotions”, psychic states based on recalled feelings, do.
The “chemo-coding” options available to the neural net involve more than (>) 10 diffusible trace metals, >90 NTs and >5 endocannabinoids , collectively >100 “dopants” (Table 1), used by the neuron to encode/decode emotive cog-info within the nECM, with combinatorially explosive encoding options [49–55].
The formation of various sets of cuinfo of varying stability presented in equation 3, is the quantal basis for the engram, the trace of memory [56, 57].
Equation 3:
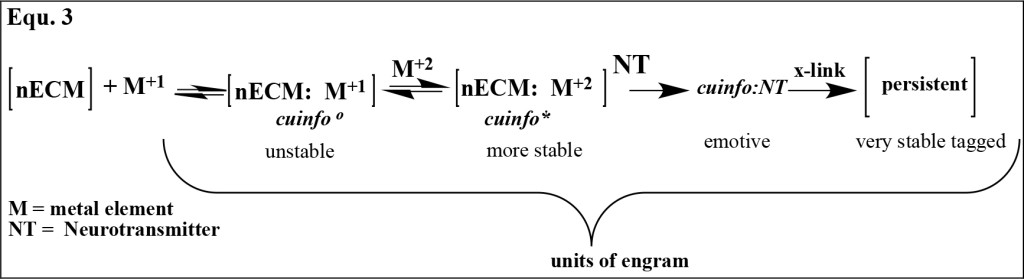
The 1st formed, original unstable cuinfo are the templates, which are “transcribed” to various anatomic compartments of the brain, where they are established as stable forms, available for decoding and consolidating into long term, persistent memory.
Discussion
The concepts of “Synaptic Plasticity” (SP) and “long term potentiation” (LTP), have been developed on the basis of Cajal’s description of the neuron and its synaptic connections to other neurons. Cajal’s model was the basis for the mathematical treatments of neural signaling by McCullogh & Pitts and adopted by Hebbs in his LTP description of neural memory. These have been adopted by the community of neurobiologists and form the basis for most current research in memory. However, serious objections were raised, the Hebbian model was accused of the 7 “sins” enumerated above.
We take a wider view of Hebb’s transgressions which stemmed from the limitations of their staining technique. Cajal used Golgi’s silver stain, which however, could not stain the nECM around neurons (see above discussion). Thus, he drew “naked neurons”, suspended in empty space. Though later generations of neurobiologists (after 1960) could no longer deny the reality of the nECM [57], they still ignored it’s possible consequences for neural signaling, as well as the “message of neural shape” (Figure 1A). Most textbooks and current articles still present images of “naked neurons” without qualifying statements about the background. Thus, later generations of neurobiologists perpetuated Cajal’s “original sin”.
Without belaboring the point, we simply state that the neuron is neither mathematical nor “naked”. But it is emotional. We propose that memory is physically encoded as a collection of cuinfo within the nECM, the neuron’s “memory material” [59–60]. The NTs could be considered the molecular coding symbols for psychic states [15–19]. Equation 3 describes the formation of sets of cuinfo with different NTs, which permit the “chemo- coding” of emotive states not available to any other coding scheme (Baudot, Braille, binary, trinary, Morse). Nor can emotive states be simulated by binary-coded algorithms [61, 62]. We suggest that the earliest formed cuinfo become the templates for those formed later, which are transduced and stored in different anatomic compartments of the brain. The consolidation of these dispersed but entangled cuinfo into seamless memory is like the cloud computing of the internet. In the interest of space, we defer a more detailed discussion of the neural “read” / “write” mechanisms for another venue.
Paradigm Shift
The acquisition of a paradigm is a sign of maturity in the development of a scientific field [63]. And exchanging one paradigm for another with greater explanatory power, signals greater maturity. One can no longer evade the aforementioned anomalies of the Cajal’s and Hebb’s model of synaptic signaling, which require atonement, a rethinking of the canons of neurobiology.
The tripartite mechanism permits one to redeem the “sins” of Cajal and Hebb, by providing a molecular rationale with 7 virtues, as follows:
7 Virtues of the tripartite mechanism
- Employs available physiologic components (neuron, nECM, metals, NTs).
- Rapid, little energy requirements (< 400 cal/day human brain).
- High (near limitless) capacity (combinatorials of Avogadro 1023 number).
- Permits the neural encoding of cog-info, with NTs as emotive signifiers.
- Describes both short and long term memory in terms of chemical stability.
- Reveals connection between memory, its loss, and inherited or drug-induced malfunctions.
- Provides a chemographic representation of cognitive units of information (cuinfo), the basic “bits” from which memory is consolidated.
Essentially, we posit that memory is physically embodied by metal-centered complexes employed by the neuron to encode, store and recall cog-info in the nECM around the neurons. This mechanism permits neural function in regard to augmented learning and memory, interpreted as SP. The tripartite mechanism of neural memory provides a context in which SP is rendered operative as for example, the functioning of “engram neurons” [62].
Conclusion
It is said that God as well as the Devil are in the details. And so too for scientists who desire molecular-scaled details of mental processes. Modern neurobiologists can redeem their “guilt” over Cajal’s “original sin” and Hebb’s lapses, by confessing that the neuron’s shape and environment are relevant to its unique mentation talent, expressed as emotive memory, stored as engrams but without Ryle’s ghosts or Augustine’s spirits
[64–66].
Hebbs assigned increased learning/memory to the phrase “Synaptic Plasticity” (SP), the improved connectivity between two neurons in synaptic contact, which are consolidated by the neural net into coherent recall (learning and remembering). Recent literature also ascribes SP to connectivity between various anatomic compartments (i.e. hippocampus, thalamus, cortex, temporal lobe, etc.) of the brain. Thus, SP has anatomic aspects as well as neural network qualities. But underneath all, lies a molecular-scaled “chemo-coded” reality which must be confessed to atone the past “sins” of neurobiology, those of ignoring the nECM and the “message” of neural shape.
The tripartite mechanism complements the observed plasticity of neural nets that become modified as a result of learning. It adds molecular definition to the talent of neural recall. In addition to clarifying the underlying function of the extended neural shape (i.e. exposure to the nECM), it identifies a coding system for emotions in the form of NTs, molecules that elicit both physical reactions and psychic states from neural creatures (Table 1) that must learn and remember to survive.
We continue to mine the rich vein of published literature to cite works which support this tripartite mechanism of neural memory with emotive qualities. In following that vein, we employ the concepts and iconography of the chemist, to propose a chemo-coding process that underlies the most obscure qualities of our being, our ability to learn and to forget.
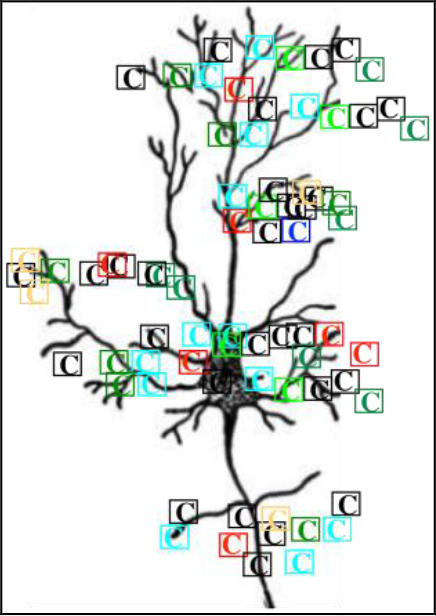
Figure 4. Schematic representation of neuron surrounded by cog-info in the form of cuinfo (![]() ) with different colors representing different NTs) formed in the nECM .
) with different colors representing different NTs) formed in the nECM .
Acknowledgement
(By GM). A memorium to my wife and fan, the artist Georgette Batlle (1940–2009), whose inspired my graphic approach to molecular reality. Thanks to friends, Lilly Rivlin (New York, N.Y.) and the late Bill Needle (Eastchester, N.Y.) for their early encouragement and financial support in the period 1980–1984. Thanks also to Karine Ahouva Leopold (Paris, Jerusalem) for many discussions on emotions and subjective states. Thanks to my brother Rabbi Dr. Tzvi Marx (Amsterdam, Jerusalem) for being a sounding board and for critical reading of the manuscript.
Conflict of Interest
GM is a founder of MX Biotech Ltd., with the commercial goal to develop new memory materials and devices.
CG is a professor emeritus of HUJI. He is active in inventing and developing of peptides and proteins-based drugs.
Not with standing, the ideas forwarded here are scientifically genuine and presented in good faith, without commercial clouding of the concepts expressed here.
References
- Hebb DO (1949) The Organization of Behavior. Wiley, New York.
- Costandi M (2017) Neuroplasticity. MIT Press, Massachusetts, USA
- Arshavsky YI (2006) ‘The seven sins’’ of the Hebbian synapse: Can the hypothesis of synaptic plasticity explain long-term memory consolidation? Progress in Neurobiology 80: 99–113.
- Willis WD Jr1 (2007) The somatosensory system, with emphasis on structures important for pain. Brain Res Rev 55: 297–313. [crossref]
- Cajal SR, Swanson N, Swanson LW (1995) Histology of the nervous system of man and vertebrates. Oxford University Press, Oxford, United Kingdom ISBN: 9780195074017.
- Vitellaro-Zuccarello L, De Biasi S, Spreafico R (1998) One hundred years of Golgi’s “perineuronal net”: history of a denied structure. Ital J Neurol Sci 19: 249–253. [crossref]
- DeFelipe J (2006) Brain plasticity and mental processes: Cajal again. Nat Rev Neurosci 7: 811–817. [crossref]
- Garcia-Lopez P, Garcia-Marin V, Freire M (2010) The histological slides and drawings of Cajal. Front Neuroanat 4: 9.
- Jones EG (2011) Cajal’s debt to Golgi. Brain Res Rev 66: 83–91. [crossref]
- Ferreira FR, Nogueira MI, Defelipe J (2014) The influence of James and Darwin on Cajal and his research into the neuron theory and evolution of the nervous system. Front Neuroanat 8: 1. [crossref]
- Switzer RC (2000) Application of silver degeneration stains for neurotoxicity testing. Toxicol Pathol 28: 70–83. [crossref]
- Bothun GD (2008) Hydrophobic silver nanoparticles trapped in lipid bilayers: Size distribution, bilayer phase behavior, and optical properties. J Nanobiotechnology 6: 13–20.
- Kandel ER (2006) In Search of Memory. WW Norton & Co New York.
- McCulloch WS, Pitts WH (1943) A logical calculus of the ideas immanent in nervous activity. Bulletin of Mathematical Biophysics 7: 115–133.
- Jeffress LA (1951) Cerebral Mechanisms in Behavior. The Hixon 1947 Symposium. Wiley, New York.
- Piccinini G (2006) Computational explanation in neuroscience. Synthese 153: 343–353.
- Körding KP, König P (2000) Learning with two sites of synaptic integration. Network 11: 25–39. [crossref]
- Frankland PW, Josselyn SA (2016) Neuroscience: In search of the memory molecule. Nature 535: 41–42. [crossref]
- Josselyn SA, Köhler S, Frankland PW (2017) Heroes of the Engram. J Neurosci 37: 4647–4657. [crossref]
- Stepanyants A, Hof PR, Chklovskii DB (2002) Geometry and structural plasticity of synaptic connectivity. Neuron 34: 275–288. [crossref]
- Ruoslahti E (1996) Brain extracellular matrix. Glycobiology 6: 489–492. [crossref]
- Dityatev A, Schachner M (2003) Extracellular matrix molecules and synaptic plasticity. Nat Rev Neurosci 4: 456–468. [crossref]
- Barros CS, Franco SJ, Müller U (2011) Extracellular matrix: functions in the nervous system. Cold Spring Harb Perspect Biol 3: 005108. [crossref]
- Kamali- Zare, Nicholson P (2013) Brain extracellular space: Geometry, matrix and physiological importance. Basic Clinical Neuroscience 4: 282–285.
- Suttkus A, Morawski M, Arendt T (2016) Protective properties of neural extracellular matrix. Mol. Neurobiol. 53: 73–82.
- Theocharis AD, Skandalis SS, Gialeli C, Karamanos NK (2016) Extracellular matrix structure. Adv Drug Deliv Rev 97: 4–27. [crossref]
- Amit D (2013) Hebb vs Biochemistry: the Fundamentalist Viewpoint.
- Bailey CH, Giustetto M, Huang YY, Hawkins RD, Kandel ER (2000) Is heterosynaptic modulation essential for stabilizing Hebbian plasticity and memory? Nature Reviews Neuroscience 1: 11–20.
- Freeman WJ (1997) Three Centuries of Category Errors in Studies of the Neural Basis of Consciousness and Intentionality. Neural Netw 10: 1175–1183. [crossref]
- Horn G (2004) Pathways of the past: the imprint of memory. Nat Rev Neurosci 5: 108–120. [crossref]
- Gallistel CR, King AP (2009) Memory and the Computational Brain. Wiley Blackwell New York.
- Anastassiou CA, Perin R, Markram H, Koch C (2011) Ephaptic coupling of cortical neurons. Nat Neurosci 14: 217–223. [crossref]
- Vizi ES, Fekete A, Karoly R, Mike A (2010) Non-synaptic receptors and transporters involved in brain functions and targets of drug treatment. Br J Pharmacol 160: 785–809. [crossref]
- Vizi ES (2013) Role of high-affinity receptors and membrane transporters in nonsynaptic communication and drug action in the central nervous system. Pharmacol Rev. 52: 63–89.
- Kandel ER, Dudai Y, Mayford MR (2014) The molecular and systems biology of memory. Cell 157: 163–186. [crossref]
- Carandini M (2012) From circuits to behavior: a bridge too far? Nat Neurosci 15: 507–509. [crossref]
- Landauer R. 1991, 1996
- Information is physical. Physics Today; May, p 93–29.
- The physical nature of information. Physics Letters A 1996; 217: 188–193
- Arbib MA (1987) Brains, Machines and Mathematics. (2ndedn). Springer Verlag, New York, USA.
- Arbib MA (2000) Warren McCulloch’s search for the logic of the nervous system. Perspect Biol Med 43: 193–216. [crossref]
- Shaik S (2003) Chemistry–a central pillar of human culture. Angew Chem Int Ed Engl 42: 3208–3215. [crossref]
- Lehninger (2008) Principles of Biochemistry. (5thedn) W.H. Freeman, New York, USA.
- Blomback M, Antovic J (2010) Essential Guide to Blood Coagulation. Wiley Blackwell, New York, USA.
- Marx G (2014) Multiple modes of blood coagulation. J Bone Marrow Res 2: 141.
- Watson J, Baker TA, Bell SA, Gann A, Levine M, et al 2013 Molecular Biology of the Gene. (7thend), ISBN-13: 978–0321762436
- Marx G, Gilon C (2016) The molecular basis of neural memory. MBM Pt 6: Chemical coding of logical and emotive modes. Int J Neurology Res. 2: 259–268.
- Marx G, Gilon C (2017) The molecular basis of neural memory. MBM Pt 7: Artificial intelligence (AI) versus neural intelligence (NI). AIMS Medical Science 4: 254–273
- Fodor JA 1975 The Language of Thought. Harvard University Press, Cambridge, MA, USA.
- Langer S (1967) Mind: An Essay on Human Feeling. Vol I, II. Johns Hopkins Press, Baltimore, USA.
- Reith, M.E 2002 Neurotransmitter Transporters: Structure, Function, and Regulation. (2ndedn), Springer Science & Business Media, New York, USA.
- Burbach JP (2010) Neuropeptides from concept to online database www.neuropeptides.nl. Eur J Pharmacol 626: 27–48. [crossref]
- Roshchin VV (2010) Evolutionary considerations of neurotransmitters in microbial, plant, and animal cells. In: M. Lyte and P.P.E. Freestone (eds.), Microbial Endocrinology: Interkingdom Signaling in Infectious Disease and Health, Springer Science & Business Media Pg No: 17–37.
- Grimmelikhuijzen CJ, Hauser F (2012) Mini-review: the evolution of neuropeptide signaling. Regul Pept 177: 6–9. [crossref]
- Agulhon C, Fiacco TA, McCarthy KD (2010) Hippocampal short- and long-term plasticity are not modulated by astrocyte Ca2+ signaling. Science 327: 1250–1254. [crossref]
- Lehn JM (1999) Dynamic combinatorial chemistry and virtual combinatorial libraries. Chem Eur J 5: 2455–2464.
- Lehn JM (2012) Constitutional dynamic chemistry: Bridge from supramolecular chemistry to adaptive chemistry. Topics Curr Chem 322: 1–32
- Schacter DL (2011) Forgotten Ideas, Neglected Pioneers: Richard Semon and the Story of Memory. Routlege, New York.
- Dudai Y (2004) The neurobiology of consolidations, or, how stable is the engram? Annu Rev Psychol 55: 51–86. [crossref]
- Kleene R, Schachner M (2004) Glycans and neural cell interactions. Nat Rev Neurosci 5: 195–208. [crossref]
- Lin WP, Liu SJ, Gong T, Zhao Q, Huang W (2014) Polymer-based resistive memory materials and devices. Adv Mater 26: 570–606. [crossref]
- Di Ventra M, Pershin YY (2011) Memory materials: A unifying description. Materialstoday 14: 584–591.
- Boole G (1853) The Laws of Thought. The Mathematical Theories of Logic and Probabilities. Project Gutenberg . Released: 02/2005.
- Turing A (1936) On Computable Numbers, with an application to the Entscheidungsproblem. Proc Lond Math Soc 42: 230–265.
- Kuhn TS (1970) The Structure of Scientific Revolutions. (2ndedn), University of Chicago Press, Chicago, USA.
- Tonegawa S, Liu X, Ramirez S, Redondo R (2015) Memory Engram Cells Have Come of Age. Neuron 87: 918–931. [crossref]
- Ryle G (1949) The Concept of Mind. University of Chicago Press, United Kingdom.
- Fredriksen P (2000) Augustine on God and memory. http://www.bu.edu/religion/files/pdf/Augustine-on-God-and-Memory.pdf